Una pieza de evidencia experimental en la división entre memoria de corto plazo y memoria de largo plazo
En 1998 Iván Izquierdo, Daniela M. Barros, Tadeu Mello e Souza, Marcia M. de Souza, Luciana A. Izquierdo y Jorge H. Medina publicaron correspondencia donde reportaban un estudio que dilucidaba la naturaleza separada de los mecanismos que hacen a la memoria de corto y la de largo plazo.
Introducción
Izquierdo et al. comienzan afirmando que por más de 100 años la pregunta principal sin respuesta en la investigación de memoria ha sido si la memoria de corto plazo es un paso necesario hacia la memoria de largo plazo. En este carta, entonces, ellos reportan cuatro tratamientos que bloquean la memoria de corto plazo, dejando intacta la memoria de largo plazo, mostrando así que estos sistemas de memoria están, hasta cierto punto, separados.
Método
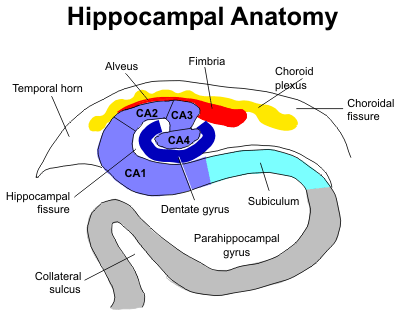
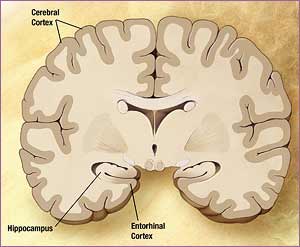
Los tratamientos que usaron los investigadores alteran todos la memoria de largo plazo cuando se inyectan en la sub-región CA1 del hipocampo o en la corteza entorrinal de ratas que han sido entrenadas para ejecutar ciertos comportamientos.
{kind=link}
{kind=link}
Estos tratamientos fueron:
- El antagonista del receptor AMPA de glutamato 6-ciano-7-nitroquinoxalina-2,3-diona (CNQX, 0,5 μg).
- El agonista del receptor GABA-A muscimol (MUS, 0,5 μg).
- El agonista del receptor 1A de serotonina 8-hidroxi-2-(di-n-propilamino) tetralin (DPAT, 2,5 μg).
- El antagonista del receptor 1A de serotonina 1-(2-metoxifenil)-4-(4-(2-phtalimido)) butilpiperazina (NAN, 2,5 μg).
Estos investigadores implantaron dos guías 1 mm por encima de la región CA1 dorsal del hipocampo o 1 mm por encima de la superficie de la corteza entorrinal de ratas Wistar (240-300 g).
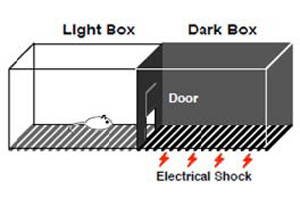
Después de la recuperación [de la cirugía], las ratas fueron puestas en una plataforma de 25 cm de alto y 7 cm de ancho. Esta plataforma estaba frente a una rejilla de acero inoxidable y fue usada para el entrenamiento de evitación inhibitoria [ver imagen de ejemplo y siguiente párrafo para ver en qué consiste].
{kind=link}
Los investigadores midieron qué tan rápido se bajaban completamente las ratas [o sea, cuánto demoraban en bajarse con las cuatro patas] hacia la rejilla. Una vez allí, las ratas recibían un choque eléctrico de 0,3 mA por 1 s en las patas. Inmediatamente después, recibían infusiones bilaterales de 0,5 μl de solución salina, vehículo (20% dimetilsulfóxido) o una droga. MUS y DPAT fueron disueltas en salina y CNQX y NAN en el vehículo.
Hubo dos experimentos principales. En el primero [intra-sujetos], los animales fueron evaluados dos veces para ver si habían retenido el recuerdo del choque eléctrico: (a) a la hora y media después del entrenamiento, para medir la memoria de corto plazo y (b) a las 24 horas después del entrenamiento, para medir la memoria de largo plazo. En el segundo experimento, Izquierdo et al. evaluaron a los animales 1,5, 3,0 y 4,5 horas después del entrenamiento.
Las sesiones de evaluación fueron similares a las de entrenamiento, con la excepción de que no hubo choque eléctrico, porque ellos estaban midiendo cuánto demoraban las ratas en bajarse a la rejilla y usaron este tiempo como una medida de su memoria del choque.
Resultados
Cuando se les proporcionó en la sub-región CA1 del hipocampo:
Discusión
Izquierdo et al. sacan varias conclusiones a partir de los resultados. En primer lugar, que los cuatro tratamientos (DPAT intra-hipocampal y CNQX, MUS y NAN intra-entorrinal) bloquean la memoria de corto plazo sin alterar la memoria de largo plazo. Esto muestra por vez primera -aseguran los investigadores- que los mecanismos de la memoria de corto plazo y los de la de largo plazo están separados.
Tercero, que receptores similares tienen diferentes roles en las dos estructuras cerebrales al mismo tiempo durante el período inmediatamente después del entrenamiento:
-En la sub-región CA1:
Conclusión
La memoria de corto plazo y la memoria de largo plazo implican, esencialmente, dos mecanismos separados.
Referencia:
Izquierdo, I., Barros, D.M., Mello e Souza, T., de Souza, M.M., Izquierdo, L.A., & Medina, J.H. (1998). Nature, 393. pp. 635-636.
Comentario
Bien, este fue el (pequeño) artículo de...hace mil años tal vez. No, mentiras, de hoy (lo que pasa es que no he podido dedicarme enteramente al blog...las ocupaciones del día a día). El caso es que en él vimos una conclusión muy simple y clara: que los mecanismos de la memoria de corto plazo son distintos de los de la memoria de largo plazo. Como vimos, también, los mismos receptores en sitios distintos pueden cumplir funciones totalmente opuestas.
Las conclusiones de esta correspondencia nos pueden parecer un tanto obvias, pero lo cierto es que fueron derivadas de evidencia experimental, hace ya 15 años. Por supuesto, no es mucho lo que podamos extrapolar a partir de este estudio, pero sí me pareció importante que lo re-conociéramos en este espacio.
¿Ideas, comentarios o sugerencias? No dudes en compartirlos con todos.
- CNQX y MUS alteraron tanto la memoria de corto como la de largo plazo.
- DPAT bloqueó la memoria de corto plazo, sin afectar la de largo plazo; el efecto en la memoria de corto plazo no fue observado más allá de tres horas después del tratamiento.
- NAN no tuvo efecto en ninguna.
Cuando se les administró en la corteza entorrinal:
- CNQX, MUS y NAN bloquearon la memoria de corto plazo sin alterar la memoria de largo plazo. El efecto sobre la memoria de corto plazo duró 3 horas.
- DPAT potenció la memoria de corto plazo y bloqueó la de largo plazo. El efecto sobre la memoria de corto plazo no se vio más allá de 3 horas después del tratamiento.
Discusión
Izquierdo et al. sacan varias conclusiones a partir de los resultados. En primer lugar, que los cuatro tratamientos (DPAT intra-hipocampal y CNQX, MUS y NAN intra-entorrinal) bloquean la memoria de corto plazo sin alterar la memoria de largo plazo. Esto muestra por vez primera -aseguran los investigadores- que los mecanismos de la memoria de corto plazo y los de la de largo plazo están separados.
En segundo lugar, Izquierdo et al. concluyen que los efectos de los tratamientos entorrinales en la memoria de corto plazo duraron más que aquellos del DPAT intra-hipocampal, lo que sugiere que los receptores intra-entorrinales "manejan" la memoria de corto plazo por un tiempo más largo.
Tercero, que receptores similares tienen diferentes roles en las dos estructuras cerebrales al mismo tiempo durante el período inmediatamente después del entrenamiento:
-En la sub-región CA1:
- los receptores AMPA son necesarios tanto para la memoria de corto como para la de largo plazo;
- los receptores GABA-A inhiben ambos tipos de memoria y
- los receptores 1A de serotonina inhiben la memoria de corto, pero no la de largo plazo.
- los receptores AMPA y 1A de serotonina son necesarios para la memoria de corto plazo;
- los receptores 1A de serotonina inhiben la memoria de largo plazo y
- los receptores GABA-A inhiben la memoria de corto plazo.
Conclusión
La memoria de corto plazo y la memoria de largo plazo implican, esencialmente, dos mecanismos separados.
Referencia:
Izquierdo, I., Barros, D.M., Mello e Souza, T., de Souza, M.M., Izquierdo, L.A., & Medina, J.H. (1998). Nature, 393. pp. 635-636.
Comentario
Bien, este fue el (pequeño) artículo de...hace mil años tal vez. No, mentiras, de hoy (lo que pasa es que no he podido dedicarme enteramente al blog...las ocupaciones del día a día). El caso es que en él vimos una conclusión muy simple y clara: que los mecanismos de la memoria de corto plazo son distintos de los de la memoria de largo plazo. Como vimos, también, los mismos receptores en sitios distintos pueden cumplir funciones totalmente opuestas.
Las conclusiones de esta correspondencia nos pueden parecer un tanto obvias, pero lo cierto es que fueron derivadas de evidencia experimental, hace ya 15 años. Por supuesto, no es mucho lo que podamos extrapolar a partir de este estudio, pero sí me pareció importante que lo re-conociéramos en este espacio.
¿Ideas, comentarios o sugerencias? No dudes en compartirlos con todos.
Comentarios
Publicar un comentario